Cell migration is an essential part of many physiological processes, such as embryogenesis1, angiogenesis2, wound healing3, and the immune response4. Unfortunately, cell migration is also involved in many pathologies, including cancer metastasis5 and inflammation6. In each of these physiological and pathological processes, the basic process of cell migration, i.e. translocation along or through a tissue substrate, is the same.
Cells migrate in different modes depending on the type and function of the cells. For example, leukocytes, for the majority of their lifespan, migrate as single cells within virtually any tissue in the body4. Many other cell types only move in specific situations to place, shape, or repair tissue. Most of these cells move in groups that are loosely or closely associated. This type of migration is called collective cell migration.
In this article, different modes of both single-cell migration and collective cell migration are discussed.
------------------------------------------------------
Researchers can easily monitor cell migration with these CytoSMART devices: full-plate scanner Omni, fluorescence cell imager Lux3 FL, mini live-cell imagers Lux2 and Lux3 BR.
------------------------------------------------------
General migration process
The basic process of cell migration starts with cells having a directional polarity, with a leading edge and a trailing end. Subsequently, actomyosin-mediated protrusions form at the leading edge followed by attachment of the protrusions to the substrate by the formation of integrin-based focal adhesions7–9. Actomyosin contraction leads to tension along the length axis of the cells, which in turn causes translocation of the cell body forward and retraction of the trailing end7–9. This basic process is relevant in most cell types and types of cell migration. However, each cell and migration type has its own specific variant (even depending on the cell environment), which can differ in terms of cell morphology, migration speed, cell-cell interactions, and dynamics7,9.
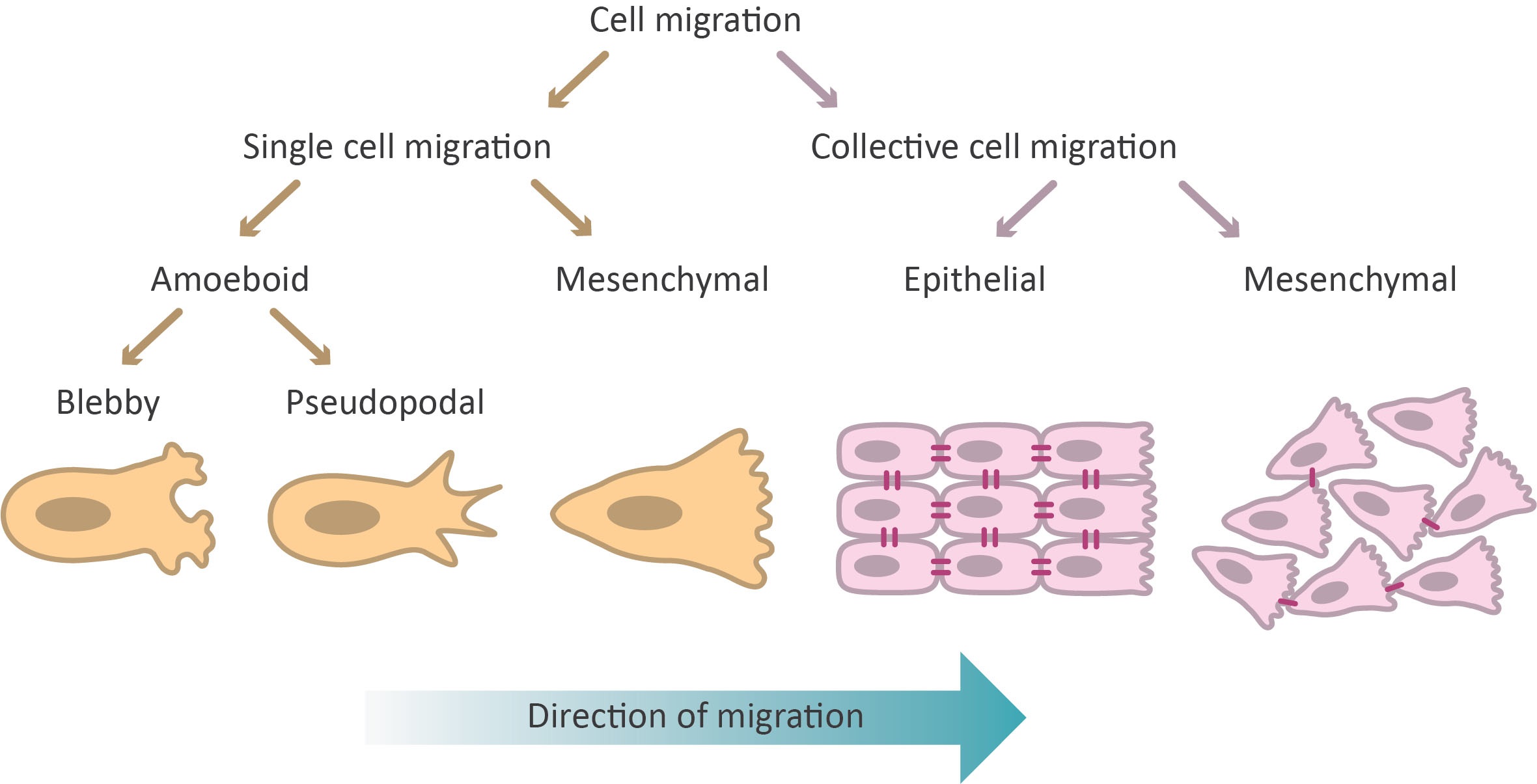
Roughly speaking, cell migration can be categorized into single-cell migration and collective cell migration. Each migration mode is then further sub-categorized into several different types of migration (Figure 1). Next to migration, cells can also display invasion. The difference between the two is that the surrounding environment is restructured by the cells in case of invasion. In the next paragraphs, single-cell migration, collective cell migration, and cell invasion are discussed in more detail.
Single-cell migration
Single-cell migration can roughly be divided into amoeboid and mesenchymal migration (Figure 1). The first form of ameboid migration is called blebby amoeboid migration and is characterized by the movement of rounded or ellipsoid cells without mature focal adhesions and filopodia7,10. These blebby cells do not migrate by adhering or pulling on the substrate but rather use propulsive, pushing blebs11. This type of migration is used by e.g. leukocytes migrating through ECM4.
Pseudopodal ameboid migration is the second form of ameboid migration and is defined by displacement of more elongated cells having weak cell-substrate interactions and actin-rich filopodia at their leading edge10,12. For example, neutrophils and dendritic cells display this type of migration13.
Elongated, spindle-like cells with strong focal adhesions and high cytoskeletal (actomyosin) contractility move using mesenchymal migration12,14. This type of migration mostly resembles the general type of migration described in section 2.1. and is seen in cells such as fibroblasts15 and sarcoma cells16.
Collective cell migration
Collective cell migration is characterized by the coordinated migration of a group of cells in which cells are influenced by interactions with each other. The exact definition of collective cell migration is still open for debate. Some argue that stable cell-cell junctions are required for collective cell migration17,18. Others suggest that migration can be considered collective when cells, moving as a group, affect each other’s movement by e.g. forming transient cell-cell contacts or secreting soluble factors9,19. Here, we comply with the latter definition and consider migration to be collective when some form of cell-cell interaction is displayed in a migrating group of cells. In that case, when looking at the extreme ends of the spectrum, collective cell migration can be split into collective migration of epithelial cells and of mesenchymal cells (Figure 1)19. However, any intermediate between epithelial and mesenchymal collective cell migration can take place depending on the cell type and its state (e.g. level of Epithelial to Mesenchymal Transition (EMT)20).
Epithelial cells form cell-cell adhesions (adherens junctions, desmosomes, tight junctions, and gap junctions) in order to fulfill their barrier function21. During collective migration, epithelial cells maintain stable cell-cell adhesions, thus still fulfilling their barrier function19,22,23. Epithelial cells can undergo collective migration in several manners. Examples of collective epithelial cell migration are the formation of sprouts or branches as seen in neo-angiogenesis of blood vessels24 and branching morphogenesis of mammary glands25. Epithelial cells can also migrate as separate groups of cells like the border cells in Drosophila egg chambers26 or invasive groups of detached cancer cells27. Another form of epithelial cell migration is the migration of strands stretching out of a tissue such as those observed in invasive carcinomas28. However, the most studied form of epithelial collective migration is that of sheet migration where cells migrate as a 2D interconnected sheet. Sheet migration is seen during wound healing, both in skin22 and in other epithelial tissues such as the intestine29 and cornea30. The common denominator between all these types of collective migration is that they rely on mechanical coupling of the cells via stable cell-cell adhesions19,22,23. These adhesions ensure coordinated cytoskeletal activity of all cells within the collective. In this manner, the group of cells can obtain a polarity and thus directionality at the collective level, similar to the polarity needed for migration at the single-cell level.
In contrast to the tightly connected epithelial cells, mesenchymal cells only form transient connections with each other. When two polarized mesenchymal cells collide, they form (N-cadherin-based) cell-cell adhesions. This triggers the retraction of the cell protrusions (lamellipodia and/or filopodia), causing a loss in polarity. This subsequently halts the migration and the cells quickly repolarize in the opposite direction. The repolarization causes the cells to move away from each other. This process is called contact-inhibition of locomotion (CIL) and can occur between cells of the same type (homotypic CIL) or between two different cell types (heterotypic CIL)31,32. Recent studies have shown that loss of heterotypic CIL is involved in cancer metastasis and invasion33–35. Next to CIL, cells actively attract each other by secreting attractants (co-attraction)19. In the case of neural crest cells, C3a is secreted, which is a well-known attractant in the immune system36. Since each cell in the group produces the same attractant, the attractant concentration is high in regions with high cell density. When a cell moves away from the group due to CIL, it can migrate back to the collective by following the local gradient of chemoattractant (e.g. C3a gradient)19. It is assumed that this continues the cycle of repulsion and attraction via respectively CIL and co-attraction maintains the collective migration of mesenchymal cells. Collective mesenchymal migration has mainly been studied in neural crest cells36–38, but recent studies show that this type of migration is also involved in many other processes, such as cancer metastasis33,39 and the migration of the mesoderm during development40.
More in-depth information about collective migration of epithelial and mesenchymal cells (including CIL) can be found in excellent reviews by Friedl and Gilmour18, Friedl and Mayor23, Rorth9, Roycroft and Mayor31, Scarpa and Mayor1, Stramer and Mayor32 and Theveneau and Mayor19.
Migration versus invasion
Next to migrating inside the body, cells can also invade their surrounding environment. In biology, the terms migration and invasion are often used as interchangeable terms. While the mechanisms are closely related, cell migration is defined as the directed translocation of cells on a 2D substrate or through a 3D matrix. Cell invasion, on the other hand, is defined as cell movement through a 3D matrix, which is accompanied by restructuring the 3D environment5,8. The process of cell invasion encompasses cell adherence to extracellular matrix (ECM) and subsequent remodeling of the ECM by means of degradation of existing ECM components and deposition of new ECM components before being able to migrate through the ECM5,8. Thus, the term invasion describes a specific mode of 3D migration, including ECM degradation, whereas migration is used to describe non-destructive movement in both 2D and 3D environments.
Related Products
There are currently no products tagged to this resource.